


The Tribalist Always Wins
Modeling "Ordo Amoris" with Evolutionary Game Theory

Introduction
On February 10th of this year, the late Pope Francis penned a rare letter directly addressing the US Council of Catholic Bishops. This letter addressing his American episcopate was not a run of the mill report or communique, it was a battle plan, an urgent call to arms in defense of Francis’ life-long devotion to a philia of immigrants. The American people had recently elected a body of leaders to revert, wholesale, the damage of the open-borders policy of migration and asylum that the Catholic Church had championed under Francis. One of those leaders is Vice President JD Vance, himself a Catholic convert, who just days prior took a torch to the progressivist conception of migrant rights. This entire letter was in response to one sentence from Vance: “Just Google ordo amoris”.

For Vance, he was repeating what he thought was the sensible, realistic, and indeed Christian conception of charity. Ordo amoris -the order of love - is an old Scholastic conception of charity, which in essence states that the cost of one's charitable acts should generally be tied to and ordered by his social obligations. A father's duty is to feed his children first, then he may care for his cousins, then he may look after his countrymen, and only then can he inquire the needs of all of humanity. This seems obvious. Francis had other ideas, and seemed to criticize Vance's conception directly:
“The human person is a subject with dignity who, through the constitutive relationship with all, especially with the poorest, can gradually mature in his identity and vocation. The true ordo amoris that must be promoted is that which we discover by meditating constantly on the parable of the “Good Samaritan” (cf. Lk 10:25-37), that is, by meditating on the love that builds a fraternity open to all, without exception.
But worrying about personal, community or national identity, apart from these considerations, easily introduces an ideological criterion that distorts social life and imposes the will of the strongest as the criterion of truth.”
Even the most casual observer of our political situation can sense that this is far more contentious than a simple spat about erroneous interpretations of Catholic social teaching. These two conceptions of social obligations are completely at odds, they are polar opposites, and the polarity defines the political situation of our time. We are all taking part in a war between the tribalist and the universal altruist, and there is no reason to suggest it couldn't be to the death.

And so, it is finally time for me to submit myself to the discussion surrounding The ChartTM, this general observation of a disorder and loss of sense of the purpose and order of altruism on behalf of our friends on the other side of the political aisle. This observation that the Left has lost its marbles when it comes to altruism and social obligations has been much discussed for a reason, and little has been left unsaid. I however will try to offer a unique insight into this observation that I have yet to see offered through evolutionary theory and population dynamics. Indeed, “Darwin cannot be cut off at the head” as it is said, and fundamental rules for evolution apply to us just as much as they do for the billions of bacterial organisms wedged under your fingernails. The Pope and the Vice President may debate to their heart's content over which is more biblical or ethical, but I am interested in answering which of the two is a winning strategy. This is the proven battleground of evolutionary theory.
Alleles and Evolutionarily Stable Strategies
Let us begin with Mendel. The term allele refers to one or more versions of a trait that fight for existence upon specific genetic loci. This is made possible in sexually-reproductive organisms due to their diploid nature: two copies of each chromosome means that mutations can produce two types of the same gene within a single organism. You may recall some of those little Punnett Squares you filled out in high school, predicting the likelihood of inheritance and expression of alleles depending on the dominant or recessive nature of the alleles. Alleles are not always static in their inheritance and expression. In fact, the very definition of evolution is the change of the frequency of alleles within a population over time. If an allele changed the reproductive fitness of an individual relative to the population mean, then the next generation will see an increased frequency of his allele. The important consideration is to conceive of two traits in evolutionary competition with one another, direct or indirect as that may be.
Tribalism and universalism can be thought of two versions, alleles, of a single trait: altruism. We will define this as any act which lowers an individual's reproductive fitness while increasing the recipient's fitness. The universalist is an altruist to all members of his species, giving freely where he can without discrimination. The tribalist on the other hand takes freely from universalists, hesitating to give back, and favoring altruism towards his own. From this conceptualization, we have several tools and theories available at our disposal to answer this question of who wins in this competition of altruistic variants. In his short article 12 Misunderstandings of Kin Selection, Richard Dawkins leans on the game theories of evolutionary biologist John Maynard Smith and gives us a framework of a model to evaluate the relative fitness of these two alleles:
“Let there be two strategies, Universal Altruist U, and Kin Altruist K. U individuals care for any member of the species indiscriminately. K individuals care for close kin only. In both cases, the caring behaviour costs the altruist something in terms of his personal survival chances. […] Every time a rare K individual behaves altruistically, it is especially likely to benefit another K individual rather than a U individual. U individuals, on the other hand, give out altruism to K individuals and U individuals indiscriminately, since the defining characteristic of U behaviour is that it is indiscriminate. Therefore K genes are bound to spread through the population at the expense of U genes. Universal altruism is not evolutionarily stable against kin altruism. Even if we assume it to be initially common, it will not remain common.”
Richard Dawkins (1979)
Dawkins here uses a specific concept in evolutionary biology developed by John Maynard Smith, an evolutionarily stable strategy (ESS). For those familiar with game theory generally, this concept is analogous to the Nash equilibrium. A strategy is stable if it is found to be “resistant to invasion”, that is, it reaches a frequency in the population where it cannot be driven to extinction by an alternative allele. Even as “players” (or genes) attempt to work around the strategies of competitors and scrape for any advantage possible, a stable strategy will find itself comfortably hull-down at a specific frequency.
The best known example of an ESS is found in sex ratios, the proportion of males to females in populations. For a long time, it was not obvious why sexually-reproductive populations all settled upon a sex ratio of roughly 1:1 - in fact, since males are observed to have higher reproductive fitness than females generally, one would expect all populations to gravitate towards a male-dominated ratio. The breakthrough in understanding why they do not is that the benefit of being a male in a population is increasingly lost, and its cost increasingly incurred, as the ratio of males increases. More competition, less mates. As some males are inevitably punished by this ratio, the ratio oscillates until it reaches equilibrium: 1:1. This is known as Fisher’s Principle. The important consideration to keep in mind moving forward is that the benefits of a trait cannot be looked at in isolation, but in context of the prevalence of other traits within the same population and how they interact with one another.
While one can see the logic in Dawkins’ reasoning pertaining to the stability (or lack thereof) for tribalist and universalist altruism alleles, he leaves a few things undefined. What constitutes “remaining common”, and what frequency would this be, exactly? As one allele trends towards complete saturation, what happens to the costs and benefits of the other? Is one driven to extinction utterly, or just low frequency? Luckily, we can use a few mathematical tricks to model out exactly what these situations look like.
Modeling Altruistic Allele Frequency

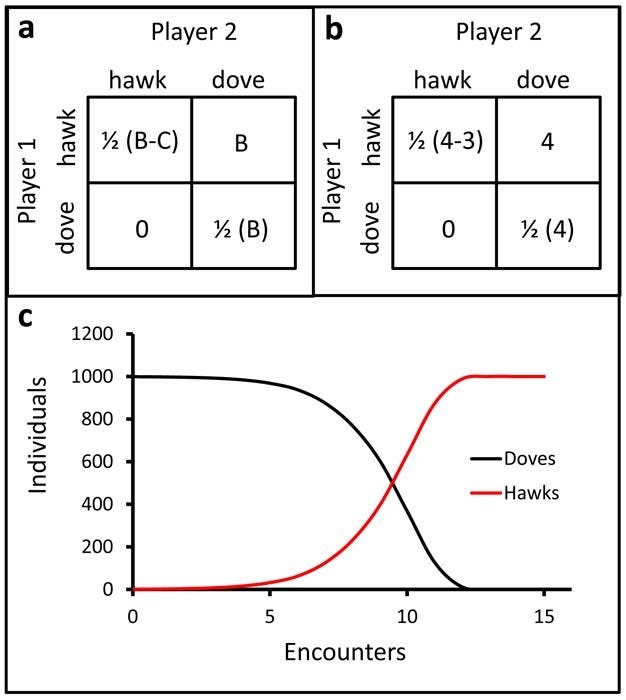
I will use Maynard-Smith’s work as a framework to make an overly simplistic model for ESS. We create a set of terms that dictate the costs and benefits to individuals possessing these alleles as they interact with one another. Maynard Smith refers to this as a payoff matrix, and we can make one of our own to lay out the expected result of a typical interaction. We will represent the tribal altruist allele with AT, and the universal altruist with AU. As suggested prior, the universal altruist gives to any member of the population, while the tribal altruist “hesitates” to give back.
In this model, we will simulate “hesitancy” through something akin to ordo amoris: when a tribalist gives to a universalist, he does so at a reduced intensity. When he gives to a fellow tribalist, he gives a full cost and receives a full benefit. This simulates the situation where a tribalist, for example, gives 75% of his wealth to his kin before giving the remaining 25% to a universalist.
There are therefore four possible interactions: a universalist gives to a tribalist (AU x AT), a tribalist gives to a universalist (AT x AU), and a interactions between two individuals of the same allele-group (AU x AU), (AT x AT). When allele-groups share with another (AT x AT) and (AU x AU), we can assume that in any random interaction a member is as likely to give as he is to receive, and at full and equivalent cost & benefit. A typical cost-benefit equation can therefore be used: payoff = benefit - cost (p = b-c). Things are a little different when these two groups interact with their competitors. When a universal altruist gives to a tribalist, he gives the full cost of altruism c while receiving a reduced benefit b’ from the tribalist. Conversely, when a tribalist gives to a universalist, he incurs a reduced cost c’ on himself and the universalist receives a full benefit b. Just for mathematical convenience, we can say that our “reduction” in cost/benefit in b’ or c’ is modified by a scaling factor k for both cases, which can be anything between 0 and 1. This way, the reduction consistently scales with the intensity of any given interaction. This gives us the following payoff matrix:
Since we are concerned with the change in relative allele frequency over time, we have to account for the change in the benefit of each strategy as its prevalence within the population changes. For example, a strategy for group cooperation would be more beneficial in a population made up of 90% group-cooperatives as opposed to a population of 10%. As evolution occurs, we expect to see the relative frequency of each change over time: one allele increases in frequency, the other decreases. We can account for this using the Hardy-Weinberg Principle (HWP), which models our universalist and tribalist alleles as such:
The expected payoffs for each strategy are calculated as the weighted average of payoffs based on the probability of meeting each type of opponent. In evolutionary terms, we can define “payoff” as reproductive fitness w, the number of offspring an individual has over an indefinite number of generations. The fitness of each allele is therefore the sum of all of its interactions with any random allele within the population, dependent upon the likelihood of meeting a specific allele. Further, since we’ve established that q = 1 - p, we can state each as:
Since we are looking at an interaction of these two alleles relative to the population’s average at any given point, fitness needs to be restated as relative to the population average w̄. Further, we need to state the frequency of the changes that results from the relative fitness of one trait being higher than the other. For each of these, we can skip a number of painful steps and adopt the replicator equation used by Maynard Smith. From there, we can plug in our own matrix and variables, such as x for our tribalist trait p:
At last, we have an equation that allows us to simulate an environment where our two altruists compete for dominance. The equation is currently termed for p, or tribalist AT, however, the inverse value for q (universalist AU) is easily obtained at each point by q = 1 - p. Now, for our parameters. Let’s assume that the altruistic act is marginally beneficial in the long run, perhaps in a reciprocal manner. The cost shall be 1, and the benefit shall be 1.25. The scaling factor k will be 0.25, signifying that a tribalist will give 5% to a universalist what he normally would a fellow tribalist. In keeping with the analysis of ordo amoris, I am using this number based on an average American charity of about 5% of annual income. The simulation can run indefinitely, but ours will end at 1000 generations. Each allele will start off at 50% of the population, a 1:1 ratio.
{kind=link}
So, who wins? At what point, if any, does the relative frequency change who benefits from the interactions? Is either an evolutionarily stable strategy, and if so, at what frequency?

Here are the results:
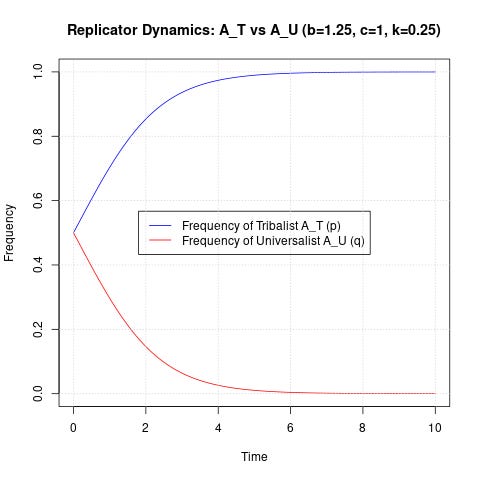
Uh oh. Uhhh….
Let’s try to mess with the numbers a little bit to see if we can produce something that isn’t immediate, catastrophic extinction for the universalists. Let’s start off universalists at 99.99% of the population, and make the scaling factor 0.9 - indicating that tribalists are only 10% less altruistic to universalists than normal:
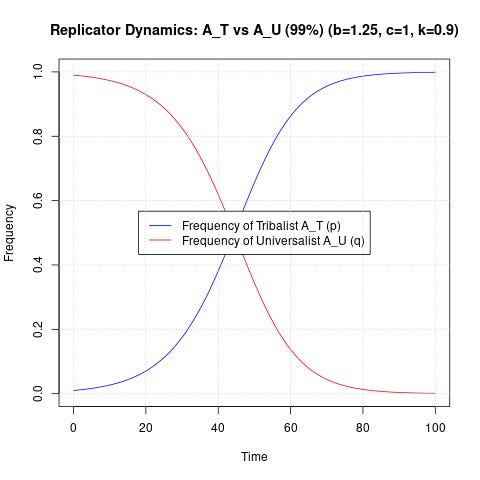
Have we made ourselves clear yet? The tribalist always wins.
Discussion
As it turns out, Dawkins was correct in his assessment of tribalist variants of altruism alleles outcompeting universalist variants. He was doubly correct in stating that the universalist allele is subject to invasion: the moment the tribalist allele enters the population, the frequency of the universalist allele plummets due to being outcompeted in terms of reproductive fitness. Wherever universal altruism is found, tribal altruism is bound to kill it. This, as Dawkins proposed, is one reason why altruism evolved precisely upon the basis of inclusive fitness. It is the evolutionarily stable strategy, resistant to invasion from competing alleles. Universal altruism, giving to any member of the population purely because they belong to the population, even if that act is ultimately beneficial to you and the recipient, is not. It is not as beneficial as tribalism, even in relatively muted forms such as that of ordo amoris.
Looking back at the model, it is obvious where this strategy goes wrong and what can be done to fix it. When a universalist meets a tribalist, the equation is entirely one-sided. The universalist gives freely, and the tribalist takes freely, but the latter gives back at a significantly reduced rate. Therefore, any and all acts of altruism towards a tribalist is costly for a universalist. For the universalist to win, he must make the act of taking more than one gives costly. In other words, he has to ruthlessly punish tribalists. He has to favor interactions with fellow universalists. He has to give deference and deterrence to groups that will help him and will harm him at indefinite timescales. In other words, the universalist must become a tribalist of his own. That, or he goes extinct.
While I won’t model the whole thing out again, I can point to a variety of studies and observations that have found exactly this. Chimpanzees will alert other males of an individual’s misbehavior, triggering a primitive justice response. All apes often become violent when sexual advances are rejected by a female, and even more violent when infidelity is afoot. All of these things are of the same bunch and what I am referring to: punishment, retaliation, and negative reciprocity.
In social animals, retaliatory aggression is common, individuals often punish other group members that infringe their interests, and punishment can cause subordinates to desist from behaviour likely to reduce the fitness of dominant animals. Punishing strategies are used to establish and maintain dominance relationships, to discourage parasites and cheats, to discipline offspring or prospective sexual partners and to maintain cooperative behaviour.
Clutton-Brock & Parker (1995). Punishment in Animal Societies. Nature. 373. 209-16. 10.1038/373209a0.
It is precisely this unwillingness to actually punish people where everything has seriously gone off the rails for the universal altruist, and why our model doesn’t have a punishment mechanism. They do not have one, not in this sense. I recall a recent discussion involving your usual “woke right” peddlers such as Seth Dillon and Joel Berry on “the dangers of the evils of tribalism”. These two no doubt had the mindset of a universal altruist in mind, invoking the Good Samaritan as Francis had. Tribalism, they lectured, cannot be responded to with more tribalism. We must rise above such rabble to a higher ideal! I hope this model sets the frame of why such a mindset is completely dead on arrival. Will such people allow us to ruthlessly punish those tribalists who favor their own group, or is that very categorization a form of naughty tribalism? They are stuck in this moralizer’s paradox of their own making and they can’t seem to get out. But I can: either we adopt an in-group preference of some kind, either we punish groups who are in fact far more tribal than we, or we die off. You may exist now, but you won’t at t = 1000. It’s just a matter of time, and we shouldn’t expect any other result.
In conclusion, we have a general and simplistic model that shows us something we already knew. Tribalists, who are selective in their charity and take care of their own first, are superior to universal altruists in terms of reproductive fitness to such an extent that the latter is doomed to extinction at the very introduction of the former. Competition with nature is rarely the first concern, it is competition within a population that poses the most serious danger to one's existence. Laws, rituals, social mores—all of these things are extensions of this observation.
The tribalist always wins.

S tier article for the thumbnail alone 🙌🔥
"The universalist is an altruist to all members of his species, giving freely where he can without discrimination." Some of them extend beyond species, not just to animals, but to barely-sentient ones like shrimp and bugs. There are utilitarians out there who claim we should devote most of our resources to other species if that's where their calculation leads.