


Epigenetics and "Blood Memory"
A deep dive into Lamarck, inheritance, and molecular biology... can epigenetics allow us to "recall past lives"?

“I’m sorry to say this because I know this is a dirty word amongst most evolutionary biologists, but — Lamarck is back! Very simple.”
-Physiologist Denis Noble, responded to by a slow and grimacing applause from the audience
Some time ago, I had a conversation with my wife on the idea of a literal interpretation of the saying, “you are your ancestors”. While she had genuine reservations about the implications this has for one's individuality, or “soul”, she did turn and tell me that often, what feels like a distant memory from some distant life would wash over her. These weren’t necessarily vivid visions, but somewhat sub-sensual experiences. She said she could almost feel and hear the winds of the Scottish highlands, and snugly-blanketed child in her arms. It was a difficult experience to explain, but I believe I understood it, because many of us have had these experiences, too.
Belief in ancestral reincarnation and the potential transmission of memories across death's firm line is one of mankind's oldest perennial convictions. This necklace from classical Greece holds a capsule containing an Orphic inscription, cautioning its wearer to avoid the River Lethe, whose waters of forgetfulness wipe clean the memories of one's life; and to instead find a pond beneath a nearby poplar tree, which will allow him to retain their memories in the next life. In the various lays of Helgi found in the Eddas, he and his lover are repeatedly killed and reborn, reuniting again and again, recalling their relationship each time. Even General Patton believed he had access to the memories of previous lives, writing in a poem attributed to him: “So as through a glass and darkly, the age-long strife I see, where I fought in many guises, many names, but always me.”

The scientific revolution has left little doubt that these ancient superstitions were simply that—superstitions. Men were not souls piloting a created body; rather, they are highly evolved animals, distinguished from nature only by degree, not by kind. Memories are but chemicals and cellular patterns, not immaterial books existing in some celestial library. The individual, as pointless and accidental as he may be, reigns supreme and requires no counsel from the past.
Yet, despite my boldness in addressing this topic, there is a certain unavoidable truth about the situation: scientific materialism is here to stay and remains the dominant method of evaluating truth. Perhaps there is a way to "prove" blood memory through other, more traditional means, be it theology or metaphysics. Maybe — I question the feasibility of such an approach so abstract. What I do know is that the findings of the 19th and 20th centuries did not keep themselves in stasis, and that today, there is compelling evidence indicating that the ancients were on to something. We may be able to uncover the biological mechanisms responsible for this phenomenon.
The ability to recall the dangers and exploits of previous lives, even if only faintly, would greatly benefit an individual’s reproductive fitness and thus be favored by natural selection. While this may seem evident, the pertinent question is not whether such a trait should evolve, but rather whether it has actually done so. What remains unclear is the mechanism behind this process. How is a memory transmitted from one generation to the next, if at all? What is the nature of that memory? Most importantly, how well does this concept align with the established modern synthesis of genetics and evolution?
This article seeks to explore these questions. It is apparent to me that blood memory indeed does exist, but only if you are willing to play with the meaning of “memory” to some degree. The key lies in an old debate in evolution, revitalized by a revolution in genetics research.
Darwinism and Lamarckism
Introductory textbooks or lectures on evolution typically begin by contrasting natural selection to an obsolete theory of evolution, Jean-Baptiste Lamarck's inheritance of acquired characteristics. In a well-known simplified example, which I am unsure if Lamarck ever even used, a blacksmith who toils away at the anvil his whole life will pass on the acquired characteristic of muscular strength to his children. In our case this theory is of great interest, because if strength can be acquired and inherited, so too can memories. Lamarck explains:
“All the acquisitions or losses wrought by nature on individuals, through the influence of the environment in which their race has long been placed, and hence through the influence of the predominant use or permanent disuse of any organ; all these are preserved by reproduction to the new individuals which arise, provided that the acquired modifications are common to both sexes, or at least to the individuals which produce the young.”
- Lamarck, Philosophie Zoologique, Ch. 7
Now, as easy as it is to pick on Lamarck for something that seems so easy to falsify today, we have to understand the context of his rationale. Firstly, Lamarck wasn’t an idiot—his model was the first true and complete theory of evolution ever put to paper, 58 years before On the Origin of Species. And in it, he correctly set the scene for the coming revolution in natural sciences: Life was not a static artifact of ancient creation. It is not separated into irreducible forms. Lamarck was the first to show that all life is connected, and in a continuous state of change, evolving. Secondly, the idea of inherited acquired characteristics was a virtual footnote in a larger model to help explain that life adapts to its surroundings, which was easily observed, but incredibly difficult to ascribe a mechanism to. He also described the idea of the use and disuse of traits in evolution, where useless traits (such as a human appendix) are gradually lost—an observation which is still useful to the theory of evolution today.
As transformational as Lamarck's theories were and as undeserving he is of the label of “the guy who got it wrong” in biology coursework, he was still of course incorrect regarding the mechanism of inherited acquired characteristics. The desires and life-actions of organisms have no impact on heritable traits, because acquired characteristics simply are not heritable. And thank God for that!—a lot of us would have been born without foreskins.
Two experiments, one famous and one infamous, finally put this idea to bed. In 1888, August Weismann distinguished somatic cells from germline cells, and more importantly the observation that genetic material could not be passed between them—later dubbed the Weismann Barrier. In other words, mutations to the body are not passed on. His idea to test this in relation to the inheritance of acquired characteristics was quite simple: breed a population of mice, and cut off their tails each generation. After 22 generations and not one newborn mouse with a short tail, he ended the experiment satisfied that the inheritance of acquired characteristics was not how organisms adapted to their environment.
Just a few decades later, an Austrian zoologist named Paul Kammerer believed he had uncovered experimental evidence to prove Weismann a fool through a series of breeding experiments involving the model organism Alytes obstetricans, the midwife toad. Kammerer wanted to show the inheritance of acquired characteristics through nuptial pads on male toads, forearm growths which assist the male in gripping females during mating in wet conditions. In a controlled environment, Kammerer simply cranked the heat up in the enclosure to force the normally dry-land-loving toads into the cooler water. He hypothesized that in these wet conditions, the nuptial pads will gradually increase in size in response to the environment over several generations, just as Lamarck had hypothesized. And in his experiments, indeed they did. From 1906 to 1926, a serious debate raged in biology over these nuptial pads. Kammerer enjoyed fame and an academic tour which greatly alleviated his financial troubles. Yet, that abruptly ended when Gladwyn Noble wrote a letter to Nature stating that he had acquired one of Kammerer’s toads, and that on these nuptial pads were injection sites filled with black ink. A serious scandal had blown its lid. Weeks later, Kammerer’s body, a gun, and a suicide note pleading that he had been framed were discovered on a mountaintop above nearby Vienna.

And that was that. The fall of Kammerer went in tandem with the final fall of Lamarckism. Darwinism was no longer competing with other theories on the periphery, and it became the undisputed creed of evolutionary science. With the discovery of DNA and its role in storing hereditary information soon after, Darwin’s model was finally complete with a molecular mechanism of inheritance. The Modern Synthesis of Darwin and Mendel was born, and this has been our understanding of evolution ever since.
That puts a nip in the bud for any material argument for “blood memory”, rather conclusively. If acquired characteristics cannot be passed down, then learned experiences, acquired memories, cannot be written into the genetic structure of germ cells and passed on to our descendants. Case closed.
Or is it?
New developments in genetics have led us to conjure up the ghosts of Lamarck and Kammerer. We will see if these revelations have any validity or use for our discussion, but first, we must zoom in to the level of the gene.
Genes, Prions, and Immortal Replicators
It is essential to understand what genes are for and what they’re doing at the molecular level for our discussion. The “central dogma of molecular biology” is fairly well known and understood: DNA transcribes RNA, and RNA makes proteins, so described by James Watson, co-discoverer of the double-helix. This unidirectional pathway from gene to protein means that the phenotype of an organism, or more generally what it is, is set in stone at the moment of conception. The environment is therefore unable to actually change the written code of the individual's genome at any heritable level and drive evolution. Of course, this is somewhat of a generalization. The inner mechanisms of the eukaryotic cell are indecipherably complex, and while life is forced to abide by the laws of nature, it also seeks to push and take advantage of those laws wherever possible. For example, hereditary information can be cycled backwards from RNA to DNA in a process known as reverse transcription. This led to Watson's peer Francis Crick to reframe the position:
“The central dogma of molecular biology deals with the detailed residue-by-residue transfer of sequential information. It states that such information cannot be transferred from protein to either protein or nucleic acid.”
- Crick, Nature, 1970

In sum, the sequence of hereditary information, once it is translated into a protein, cannot be transformed back into DNA, or RNA, or some other protein. This is something that only sequences of residues, or nucleotides, can do. This is why evolution is fundamentally gene-centric, meaning that since the function and production of cellular proteins occurs only through the structure and makeup of genes, the change in the frequencies of phenotypes (caused by proteins) within a population over time—evolution—occurs precisely at the level of genes.
But an interesting quirk can be seen in the example of prion propagation. Prions are, to use a simplistic comparison, a protein-level virus, misfolded proteins which “infect” other proteins by incurring them to conform to the misfolded shape. In mammals, the PRNP gene encodes for PrPC, a monomeric protein which performs a number of essential duties for neurons. If the infectious, misfolded variant PrPSc is introduced to the cell, the variant binds with PrPC and conforms it into the shape of PrPSc through a process called seeded polymerization. From here, the rate of infection is exponential, until the cell can no longer function. The result is a terrifying, exponential intensification of neuro-degenerative symptoms caused by mass cell death in nervous tissue.
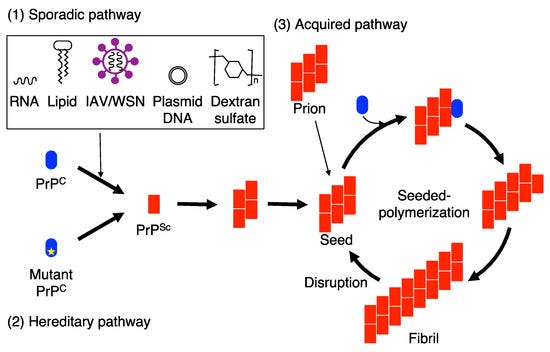
Since proteins lack genetic information and the Central Dogma forbids any retroactive transfer of the information that made them, it is more accurate to call this infection “propagation” rather than “reproduction”. But still, we do observe a form of replication occurring here, perhaps accurately called “analog replication” as opposed to the traditional “digital replication” of DNA/RNA.
Once this bridge is crossed, and indeed it has, one quickly notices the muddied picture that the Central Dogma once provided more clearly, as well as the implications for evolution at the molecular level more generally. For starters, if something can replicate, then forms of itself which replicate better will eventually become the dominant variant. In other words, it can evolve. Remember, there are no nucleotides at play here at all—only proteins. For example, PrPSc is highly resistant to protease enzymes, which exist to degrade and destroy mutated proteins that threaten the cell. This is an artifact of selection, as any variant susceptible to protein degradation would be an inferior replicator to variants which resist it. This conception of a replicator without, or before, DNA/RNA has been hypothesized for some time, including in theoretical models of abiogenesis through autocatalytic sets, and Richard Dawkins in The Selfish Gene:
“Replicators are no longer peppered freely through the sea; they are packaged in huge colonies—individual bodies. And phenotypic consequences, instead of being evenly distributed throughout the world, have in many cases congealed into those same bodies. But the individual body, so familiar to us on our planet, did not have to exist. The only kind of entity that has to exist in order for life to arise, anywhere in the universe, is the immortal replicator.”
- Dawkins, The Selfish Gene, Ch. 13
Not only do we have a curious case of evolution and replication being driven outside of genes—indeed, epigenetically—we also have an evident case of prion proteins changing the very gene expression of their unfortunate host, and in more direct ways than simple natural selection of the resistant. For example, the PSI+ prion, a misfolded variant of the translation release factor Sup35p, has been observed in yeast cells. With this protein replaced by a malfunctioning variant, stop codons are no longer able to be read during protein synthesis. In turn, portions of the genome that have been suppressed or silenced by stop codons are now active, and previously hidden genes are being transcribed. What phenotypic changes occur as a result of this depends on what comes after the stop codon, but the potential resulting variation could expand the cell’s entire genome. The prion has, in effect, changed the gene expression of the cell at random.
However, these proteins are not changing the genome, as Crick had accurately predicted: such a pathway simply does not exist anywhere in the toolkit of life on Earth. But it is, in a very primitive way, driving evolution through heritable means. Though the Central Dogma remains technically correct, it may have rather missed the point, as we are seeing heritable changes in the variances of phenotype occurring entirely outside of genes themselves.
The prion is a specific example of a larger phenomenon of extra-genetic, or rather epigenetic, changes to the hereditary information of an organism. The Weismann Barrier, The Central Dogma— none of it is a firm exclusion in this instance as previously believed: though genes are involved throughout, these changes do not require changes to the actual residue sequence of the genome to incur phenotypic variation, and indeed, the passing on of traits from a parent organism to its offspring… perhaps even memory.
Epigenetics, Unpackaged
We have finally arrived at the key buzzword of this article: epigenetics. As previously discussed, the potential for heritable memory depends on a mechanism that can effectively circumvent the Weismann Barrier and the Central Dogma, so that an “acquired” trait such as memory can be passed on to the next generation. Epigenetics promises exactly as much. But how?
Let us start back with the typical, every-nano-second case of DNA transcription. To make a protein, a cell must first transcribe a DNA sequence into an RNA sequence, which will then be translated into a protein. The process of DNA transcription is an incredibly complex miracle of life, involving hundreds of different types of polymerases, coenzymes, transcription factors, signal proteins, termination factors… an innumerable amount of molecules in an unimaginable web of interactions. To get the job done, this set of molecular machinery has to be able to freely access regions of DNA. But there is one small problem: over the course of evolutionary history, the human genome has somehow packaged nearly 2 meters of lines of double helix in each and every last one of its 10 trillion individual cells. The cell therefore requires not only an incredibly efficient means of storage, but also even more machinery to unpack specific segments when necessary.
The cell’s way of accomplishing this is a series of molecular packaging. Strands of DNA are tightly wound up around histone proteins, which themselves are wound into nucleosomes, those into chromatin, and finally into chromosomes. The end result is billions of nucleotide pairs in 2-meter-long strands neatly packed into chromosomes no more than a few micrometers in length.


Now, all the cell needs to figure out is how to actually access the 3 billion base pairs it just shoved into a dense pad of steel wool. More importantly, it needs to be able to access specific segments individually at the precise moment those segments are needed. Luckily, it has evolved yet another system of machinery for this exact task: the epigenome. The strands of DNA didn’t just “do that”, they were pushed and pulled by a system of proteins, signals, and molecules which dictate when they wrap up or unfold. Attached to the ends of the histone “beads” which the DNA “string” wraps around are histone tails, which terminate with binding sites for several potential post-translation modifications. The two most important for this topic are methylation and acetylation. If a methyl group attaches to a histone tail, the chemical structure of the histones close in, compacting the nucleosome and restricting transcription. If an acetyl group attaches, the opposite occurs, the nucleosome relaxes in structure and transcription is enabled.
Remember, you and I started off as a single-celled zygote, yet within that individual cell contained all of the genetic information necessary to differentiate into bones, neurons, glial cells, and Leydig cells. It is this packing-and-unpacking of genetic material that allows for such a diverse array of differentiated cells, all with the same genes. Just add a methyl or acetyl group, simple as that! Well, the eerie part comes with how the cell determines when, where, and how to put these groups. Enter another orchestra of molecules… The enzymes DNA methyltransferase (DNMT), histone acetyltransferase (HAT), histone methyltransferase (HMT), histone deacetylase (HDAC); the signal proteins: growth factors, cytokines, SMADs… on and on. To demystify an admittedly mystical arrangement, the cell is using a wide toolkit to respond to its external chemical environment. A specific environment prompts a specific response from the cell, and it has seen many environments over its 4-billion-year evolutionary history.

For example, a small gene on Chromosome 5 by the name of NR3C1 codes for the glucocorticoid receptor (GR) protein, the cell’s way of responding to levels of cortisol in its chemical environment. In response to stress hormones, GR moves into the nucleus and becomes a transcription factor, signaling to the cell’s transcription machinery to produce more anti-inflammatory proteins. But something very interesting happens in high levels of chronic stress. In a chemical environment saturated with cortisol, the cell naturally seeks to reduce its sensitivity to cortisol and return to a stable state. Signaling cues from the hypothalamic-pituitary-adrenal complex (HPA) direct DNA methyltransferases to the NR3C1 loci, which slaps a methyl group onto the tail of a nearby histone, silencing the gene and the production of more GR proteins. In textbooks, this mechanism is described using the example of rats licking their pups, which incurs this modification of the NR3C1 gene.
Simple enough. The cell is using highly-evolved machinery to respond to its chemical environment. The genes make the machinery, the machinery regulates the genes, and a beautiful dance between all parties ensues to strive towards survival. But let’s throw one more wrench into the equation. Not only is the NR3C1 gene heritable, of course, but so too is its methylated state. Epigenetics is heritable, across multiple generations.
Transgenerational Epigenetic Inheritance
Now, we have a clear toolkit to explain how acquired traits can be heritable without interfering with clear rules of biology. In essence, cellular responses to its chemical environment triggers a signaling cascade that structures chromatin in such a way that it can access genes appropriate for that environment. Since this structuring is heritable, the gene expression that comes as a result of it is passed down to multiple future generations. This specific phenomenon is termed transgenerational epigenetic inheritance (TEI).
Several notable experiments on model organisms, similar to that of Kammerer’s, have shown evidence of TEI and the persistence of inherited memory across multiple generations. One sticks out: a recent controversial paper in Nature tested TEI by exposing male rats to a lovely scent of sweet cherries, followed immediately by a painful shock. After some time, the cherry scent prompted a visible fearful response in the rats, including shaking and wincing. The researchers then took their sperm, created a new generation through in-vitro fertilization, and put the pups in a new container. After leaving them alone and in isolation from their parents, they reintroduced the cherry scent that the new generation had never smelled. They winced and shook, anticipating a shock they had never felt. The experiment was repeated, this time skipping a generation entirely, and the phenomenon persisted. Seeking to explain the phenomenon, they looked at olfactory receptor genes to search for changes, and indeed, the OLFR151 genes in the germ cells of these rats were littered with methylation markers.
Though it is incredibly difficult to study at-scale for humans, evidence is growing for TEI in our own lives and development. Recall the NR3C1 gene from earlier and its role as a response to stress. In postmortem analyses of brain tissue of suicide victims, those who had experienced childhood abuse and/or were descendants of people who themselves experienced it, had methylation markers on the gene. The Överkalix Cohort Studies showed that grandparents exposed to low or high stress environments influenced the health of their grandchildren through potentially epigenetic means.
How exactly somatic cells successfully signaled to germ cells this information and incurred a response to the stress is not known. A number of complex theories are being suggested, including long-living RNA strands that travel in the bloodstream, but no one knows for sure. What we do know is that the acquired fearful memory of scent was inherited, without parentage/rearing or any form of fetomaternal communication.
Though the molecular level isn’t clear, the evolutionary level seems far more simple. The organism has evolved on the rationale that the offspring should “anticipate” being put into the same environment as its ancestors. The epigenetic inheritance of memory could be the cell's clever way of transmitting such an anticipation across generations—much to the benefit and reproductive success of future generations, as opposed to those thrown into an unexpected environment.
Epigenetic… Blood Memory?
We return to the big question at hand with all of the necessary background information necessary to tackle it. Is it possible for memories to be passed down generations?
Due to the firm constraints of the Weismann Barrier and the Central Dogma, as well as a handful of experimental evidence, epigenetics shows itself to be the most promising candidate. The experiences of the parent are signaled to their germline, with the evolutionary cause of preparing the next generation for the environment they will find themselves in.
But there are a number of problems with this theory. Firstly, we have to remind ourselves what a memory actually is. Memories are stored in the brain of individuals through the synaptic connection of neurons, and the varied strength or weakness of that connection. In contrast, epigenetic inheritance deals with only one cell. Therefore, there has to be some way to translate the 3D synaptic pattern that defines a memory into the epigenetic pattern of DNA. No possible mechanism for this is known, and it is unlikely one exists. Secondly, it is unknown how long these acquired memories would even last. Some studies have shown inheritance to last up to the F9 generation, but we have little reason to believe it would be permanent. The entire point of epigenetics is to allow the cell to use its library to respond to a specific environment, and environments are never static. Once the environment changes, and change it will, there is no reason to believe the cell will leave that nucleosome open for the sake of tradition. Memories, if inherited this way, will eventually be lost. This was one small truth of Lamarck's theory regarding use and disuse: the eventual loss of the expression of useless genes.
We appear to have missed the forest for the trees in our analysis. Yes, many of us have stories and accounts of ancestral memory, but the underlying truth of the supernatural presentation of it is that it is a rare phenomenon. We don’t know the face of our grandparent's first date, we can’t recall the gate code for our great-grandparents tool shed. Seemingly all memories are lost at death, which is why the need for that gold Orphic necklace, and why we need campfire stories with our grandparents. There are clear biological limits on the quantity and quality of memories that could potentially be passed on through epigenetic inheritance.

The general problem with the fascination with epigenetics is that it insists too much upon a mechanism that the cell has already evolved to largely deprioritize over the immortal coils commanding it: DNA. To what extent epigenetics, prions, histone modifications, small RNAs, and so forth impact gene expression, it is largely irrelevant, because the simple truth of life as we know it is that each and every one of these things starts and ends with a nucleotide base pair. The cell did not invent epigenetics to control its genes—no, epigenetics itself is an invention of genes that they use to control the cell, just like every other biological process in existence.
Perhaps we do not need to couch the issue on a new theory of inheritance. What we know already could suffice in the right framing. One interpretation of a broader, more profound blood memory is that we are responding to environments in the same ways that our ancestors did. This does not rely on some evolutionary trick, but on the simple fact that we possess the same genes as our ancestors, waiting for the right conditions to be activated. Once we encounter these environments, we react just as they would have, with identical cellular machinery and cascades of activity. When we “know” what to do in a specific situation, it is because our genome has preserved the set of instructions necessary to deal with it, activated by a timely acetyl group for our benefit. Perhaps we have tried to give to blood memory what belongs more simply to instinct.
Conclusion
Epigenetics is a relatively novel curiosity, part of a larger renaissance of genetics research in the last 20 years. It has promised much, and hailed by many as a herald of necessary revisions to the Modern Synthesis of Darwin and genetics. And in some ways, it has. It has forced us to evaluate means of inheritance that do not strictly follow the Central Dogma's framing, and offered new ways to explain phenotypic variation. But eventually, once the fascination settles, we see that it is not yet ripe to reanimate Lamarck's corpse once more. Genes still rule the equation, dictate the activity of cells, and drive the mechanisms of inheritance.
At the beginning of this article, I foreshadowed that one may go about illustrating a biological explanation of blood memory, if they are willing to toy with what they mean by a memory. Despite the limitations, we still have clear evidence of memorized responses to environmental stimuli persisting across generations. We even have good ideas of how it’s done, narrowing down the search to specific genes and their respective proteins. But when people speak of blood memory, I think they are being a bit more descriptive than a temporarily-inherited deregulation of cortisol receptors.
I don’t mean this as a long-winded debunking of certain experiences with blood memory—certainly not my wife's! But I think it is important to correctly define and frame the concepts we are talking about. The living organism has every reason in the world to preserve the experiences of past lives, but at the same time, only a tiny fraction of life has evolved what could even begin to be called “memory”. For that reason and many others, what we believe we are sensing is likely far more primitive, far more molecular, than one would originally believe. In the end, we may not need to look further than our own genomic sequences in search of the lives of our ancestors.
Indeed, we are them.
Thanks for reading!
Further Reading
Watch this video simulating some of the cellular processes discussed. It is one of the most awe-striking videos you will ever watch.
Dawkins's The Selfish Gene is a fantastic read with less intensive language than this article uses. If you enjoy a full-dive, try The Extended Phenotype.
Various papers and studies cited throughout the article:
Colonna, Federica Turriziani, "Paul Kammerer's Experiments on the Midwife Toad (1905-1910)". Embryo Project Encyclopedia ( 2014-12-30 ). ISSN: 1940-5030 https://hdl.handle.net/10776/8279
Koonin EV. Does the central dogma still stand? Biol Direct. 2012 Aug 23;7:27. doi: 10.1186/1745-6150-7-27. PMID: 22913395; PMCID: PMC3472225.
Serio TR, Lindquist SL. [PSI+]: an epigenetic modulator of translation termination efficiency. Annu Rev Cell Dev Biol. 1999;15:661-703. doi: 10.1146/annurev.cellbio.15.1.661. PMID: 10611975.
Crick, Francis. Central Dogma of Molecular Biology. Nature 227, 561–563 (1970). https://doi.org/10.1038/227561a0
Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York: Garland Science; 2002. Chromosomal DNA and Its Packaging in the Chromatin Fiber.
Fitz-James, M.H., Cavalli, G. Molecular mechanisms of transgenerational epigenetic inheritance. Nat Rev Genet 23, 325–341 (2022). https://doi.org/10.1038/s41576-021-00438-5
Dias, B., Ressler, K. Parental olfactory experience influences behavior and neural structure in subsequent generations. Nat. Neurosci. 17, 89–96 (2014). https://doi.org/10.1038/nn.3594
Misztak P, Pańczyszyn-Trzewik P, Nowak G, Sowa-Kućma M. Epigenetic marks and their relationship with BDNF in the brain of suicide victims. PLoS One. 2020 Sep 24;15(9):e0239335. doi:10.1371/journal.pone.0239335.
Vågerö, D., Pinger, P.R., Aronsson, V. et al. Paternal grandfather’s access to food predicts all-cause and cancer mortality in grandsons. Nat Commun 9, 5124 (2018). https://doi.org/10.1038/s41467-018-07617-9
Horsthemke, B. A critical view on transgenerational epigenetic inheritance in humans. Nat Commun 9, 2973 (2018). https://doi.org/10.1038/s41467-018-05445-5
Really great post. Funnily enough, Darwin's theory of inheritance was that all your organs shed some type of particle called gemmules which gets passed on to your kids. Like a liver gives off tiny liver particles which travel to your sperm and then rearrange in your offspring to make a liver again. And using your liver more might make it give off more gemmules and make your offsprings liver bigger. Today this would be called Lamarckism!
Awesome article, really well written and important material discussed respectfully and in-depth, this might be one of the best works you've done, I admire you for covering this topic. The Patton poem was unknown to me, I simply know of a quote from him about the topic. I'll have to look for more of Patton's poetry.
One of the key reasons I returned to paganism is because I have long believed in genetic memory, blood memory, and reincarnation. I used to think of this as direct soul reincarnation, and sometimes still think of it that way, but more often than not I believe that I have memories encoded in my genes which bubble up from time to time in the form of instincts. Since the ideal and spiritual is very real to me, and the foundation of this material realm, I think that genes are likely capable of reaching over, their storage systems are quite robust as you say. Jean-Emile Charon speaks about the electron as being a sort of "cross-over" point between the material and immaterial, and he partly argues based on their incredible storage systems. There are many other things he talks about, he was primarily a physicist, and I am neither a geneticist or a physicist, nor really a theologian, I am just a farmhand and poet.
This is why I think that in a religious sense, it may be worthwhile to speak of the DNA as a miniature Tree of Life, a World-Tree within us, that which makes our bodies temples, a subliminal space within our very cells. This might bring fruit if explored.